- 現在位置
- トップ > 科学技術・学術 > 研究費、研究開発評価 > 文部科学省の競争的資金一覧 > 科学研究費補助金 > 科学研究費補助金「特別推進研究」の追跡評価(試行)について > 「特別推進研究」研究期間終了後の効果・効用、波及効果に関する自己評価書
「特別推進研究」研究期間終了後の効果・効用、波及効果に関する自己評価書
- 研究代表者氏名
竹市 雅俊(理化学研究所・高次構造形成研究グループ・グループディレクター) - 研究分担者氏名
千坂 修(京都大学・理学研究科・助教授)
上村 匡(京都大学・理学研究科・助手) - 研究課題名
「シナプス結合と神経回路の形成機構」 - 課題番号
10102006 - 補助金交付額(直接経費のみ)
平成10年度 80,000千円 平成11年度 60,000千円 平成12年度 47,000千円 平成13年度 67,000千円 平成14年度 67,000千円
1.特別推進研究の研究期間終了後、研究代表者の研究がどのように発展したか
(1)概要
研究代表者は、平成14年(2002年)4月、京都大学大学院生命科学研究科から、独立行政法人理化学研究所発生・再生科学総合研究センターに、センター長、並びに、高次構造形成研究グループのグループディレクターとして移籍し、本特別推進研究を継続した。終了後、引き続き新しい特別推進研究課題「接着装置に依存した新しい細胞行動制御シグナルの探索」が採択され(研究期間:平成15年~18年度)、これにより、前課題の中心テーマであったシナプス形成の制御に関する研究をさらに発展させると共に、シナプス形成の土台となっている細胞接着現象についてより一般的な細胞生物学的機構を解明するための研究を発展させた。また、理化学研究所の運営費交付金によって、細胞接着分子と神経発生の関係に関する研究も併せて進めた。その結果、平成14年度までの特別推進研究の成果は、その後の研究の発展のために極めて有効に生かされ、平成15年~18年度間の特別推進研究の成果の事後評価では(平成19年10月実施)では、(A![]() )の評価を得ている。
)の評価を得ている。
(2)論文発表、国際会議等への招待講演における発表など
論文発表
- Kubo, F., Takeichi, M., and Nakagawa, S. (2003) Wnt2b controls retinal cell differentiation at the ciliary marginal zone. Development 130,587-598.
- Nakagawa, S., Takada, S., Takada, R., and Takeichi, M. (2003) Identification of the laminar-inducing factor: Wnt-signal from the anterior rim induces correct laminar formation of the neural retina in vitro. Develop. Biol., 260, 414-425.
- Tanabe, K., Takeichi, M., and Nakagawa, S. (2004) Identification of a nonchordate-type classic cadherin in vertebrates: Chicken Hz cadherin is expressed in horizontal cells of the neural retina and contains a nonchordate-specific domain complex. Dev. Dynamics 229, 899-906.
- Abe, K., Chisaka, O., van Roy, F., and Takeichi, M. (2004) Stability of dendritic spines and synaptic contacts is controlled by αN-catenin. Nature Neuroscience 7, 357-363.
- Tanoue, T., and Takeichi, M. (2004) Mammalian Fat1 cadherin regulates actin dynamics and cell-cell contact. J. Cell Biol. 165, 517-528.
- Shima, Y., Kengaku, M., Hirano, T., Takeichi, M., and Uemura T. (2004) Regulation of dendritic maintenance and growth by a mammalian 7-pass transmembrane cadherin. Develop. Cell 7, 205-216.
- Nakao, S., Uemura, M., Aoki, E., Suzuki, S.T., Takeichi, M., and Hirano, S. (2005) Distribution of OL-protocadherin in axon fibers in the developing chick nervous system. Brain Res. Mol. Brain Res. 134, 294-308.
- Takeichi, M., and Abe, K. (2005) Synaptic contact dynamics controlled by cadherin and catenins. Trends in Cell Biol., 15, 216-221.
- Tanoue, T., and Takeichi, M. (2005) New insights into Fat cadherins. J. Cell Sci., 118, 2347-2353.
- Kubo, F., Takeichi, M., and Nakagawa, S. (2005) Wnt2b inhibits differentiation of retinal progenitor cells in the absence of Notch activity by downregulating the expression of proneural genes. Development 132, 2759-2770.
- Togashi, H., Miyoshi, J., Honda, T., Sakisaka, T., Takai, T., and Takeichi, M. (2006) Interneurite affinity is regulated by heterophilic nectin interactions in concert with the cadherin machinery. J. Cell Biol., 174, 141-151.
- Uemura, M., and Takeichi, M. (2006) αN-catenin deficiency causes defects in axon migration and nuclear organization in restricted regions of the mouse brain. Dev. Dynamics 235, 2559-2566.
- Otani,T., Ichii, T., Aono, S., and Takeichi, M. (2006) Cdc42 GEF Tuba regulates the junctional configuration of simple epithelial cells. J. Cell Biol., 175, 135-146.
- Tanabe, K., Takahashi,Y., Sato, Y., Kawakami, K., Takeichi, M., and Nakagawa, S. (2006) Cadherin is required for dendritic morphogenesis and synaptic terminal organization of retinal horizontal cells. Development 133, 4085-4096.
- Takeichi, M. (2006) The cadherin superfamily in neuronal recognitions and interactions. Nature Reviews Neurosciences, 8, 11-20.
- Nagae, S., Tanoue, T., and Takeichi, M. (2006) Temporal and spatial expression profiles of the Fat3 protein, a giant cadherin molecule, during mouse development. Develop. Dynamics 236, 534-543.
- Kametani, Y., and Takeichi, M. (2007) Basal-to-apical cadherin flow at cell junctions. Nature Cell Biol., 9, 92-98.
- Kadowaki, M., Nakamura, S., Machon, O., Krauss, S., Radice, G.L., and Takeichi M. (2007). N-cadherin mediates cortical organization in the mouse brain. Develop. Biol., 304, 22-33.
- Hoshino, H., Uchida, T., Otsuki T., Kawamoto, S., Okubo, K., Takeichi, M., and Chisaka, O. (2007) Cornichon-like Protein (CNIL) Facilitates Secretion of HB-EGF and Regulates Proper Development of Cranial Nerves. Mol Biol Cell. 18,1143-1152.
- Abe, K., and Takeichi, M. (2007) NMDA Receptor activation induces calpain-mediated β-catenin cleavages for triggering gene expression. Neuron 53, 387-397.
- Lee, D.M., Kiener, H.P., Agarwal, S.K., Noss, E.H., Watts, G.F., Chisaka, O., Takeichi, M., and Brenner, M.B. (2007) Cadherin-11 in synovial lining formation and pathology in arthritis. Science 315, 1006-1010.
- Suzuki, S.C., Furue, H., Koga, K., Jiang, N., Nohmi M., Shimazaki, Y., Katoh-Fukui, Y., Yokoyama, M., Yoshimura, M., and Takeichi, M. (2007) Cadherin-8 is required for the first relay synapses to receive functional inputs from primary sensory afferents for cold sensation. J. Neurosci. 27, 3466-76.
- Ichii, T., and Takeichi, M. (2007) p120-catenin regulates microtubule dynamics and cell migration in a cadherin-independent manner. Genes to Cells 12, 827-839.
- Uemura, M., Nakao, S., Suzuki, S.T., Takeichi, M., and Hirano, S. (2007) OL-protocadherin is essential for growth of striatal axons and thalamocortical projections. Nature Neuroscience, 10, 1151-1159.
国際会議での招待講演
- First Annual Symposium in Morphogenesis and Regenerative Medicine, University of Virginia (Charlottesville), May 19-21, 2003 (Invited Lecture)
- ComBio2003 (Melbourne), September 28-October 2, 2003(Plenary Lecture)
- International EMT Meeting (Port Douglass), October 5-8, 2003 (Invited Lecture)
- 27th Annual Mid-Winter Research Meeting for Association for Research in Otolaryngology (Daytona Beach), February 22-26, 2004 (Invited Lecture)
- Gordon Research Conference on Signaling by Adhesion Receptors (Bristol), June 20-25, 2004 (Plenary Lecture)
- 16th International Congress of the IFAA (Kyoto), August 22-27, 2004 (Invited Lecture)
- 2nd International DFG Symposium “Cell Migration and Disease” (Karlsruhe), October 10-13, 2004(Plenary Lecture)
- Gordon Research Conference on Cell Contact and Adhesion (Andover), June 26-July 1, 2005 (Plenary Lecture)
- ISN-ESN Innsbruck 2005 (Innsbruck), August 22-26, 2005 (Invited Lecture)
- 15th International Society of Developmental Biologists Congress 2005 (Sydney), September 3-7, 2005 (Invited Lecture)
- Toshi Yamada Memorial Lecture (Brisbane), March 15, 2006(Plenary Lecture)
- JSPS-NIH Symposium: Frontiers in 21st Century Biomedical Science (Besthesda), November 6-7, 2006 (Invited Lecture).
- Keystone Symposium “Tissue Engineering and Developmental Biology” (Snowbird), April 12-17, 2007 (Invited Lecture).
- Gordon Research Conference on Cell Contact and Adhesion (II Ciocco), May 27-June 1, 2007 (Invited Lecture).
(3)研究費の取得状況
特別推進研究(研究期間:平成15年~18年度)
研究課題名「接着装置に依存した新しい細胞行動制御シグナルの探索」
研究代表者、研究経費256,000千円
(4)特別推進研究の研究成果を背景に生み出された新たな発見・知見
1)シナプス形成におけるカドヘリン接着装置の役割
 シナプス形成のためにカドヘリン接着装置が関与することを明らかにした。その後の発展として、カドヘリン結合因子の一つαN-カテニンを欠失した海馬神経細胞の性質を詳細に解析した結果、この分子がなくともシナプスは形成されるが、その構造が非常に不安定であること、逆に、αN-カテニンを過剰発現させるとシナプスが異常に安定化すること、さらには、αN-カテニンのシナプスにおける局在が神経活動によって増減することを見出し、本分子がシナプスの安定性の調節機能を担う可能性が示唆された。
シナプス形成のためにカドヘリン接着装置が関与することを明らかにした。その後の発展として、カドヘリン結合因子の一つαN-カテニンを欠失した海馬神経細胞の性質を詳細に解析した結果、この分子がなくともシナプスは形成されるが、その構造が非常に不安定であること、逆に、αN-カテニンを過剰発現させるとシナプスが異常に安定化すること、さらには、αN-カテニンのシナプスにおける局在が神経活動によって増減することを見出し、本分子がシナプスの安定性の調節機能を担う可能性が示唆された。 カドヘリンは20種類以上あって、それぞれが別の神経細胞で発現されている。しかし、個々のカドヘリンの役割が不明であった。本特別推進研究の期間中に、これらのカドヘリンの一つcad8(カドヘリン8遺伝子)のノックアウトマウスを作製し、その後、表現型の解析を続けた。その結果、このカドヘリンが感覚神経細胞のうち、低温を感ずる細胞で発現していること、cad8(カドヘリン8遺伝子)を失ったマウスでは、低温感受性に異常が生じることなどが分かり、初めて特定のカドヘリンの生理的役割を明らかにすることができた。
カドヘリンは20種類以上あって、それぞれが別の神経細胞で発現されている。しかし、個々のカドヘリンの役割が不明であった。本特別推進研究の期間中に、これらのカドヘリンの一つcad8(カドヘリン8遺伝子)のノックアウトマウスを作製し、その後、表現型の解析を続けた。その結果、このカドヘリンが感覚神経細胞のうち、低温を感ずる細胞で発現していること、cad8(カドヘリン8遺伝子)を失ったマウスでは、低温感受性に異常が生じることなどが分かり、初めて特定のカドヘリンの生理的役割を明らかにすることができた。 カドヘリンがシナプス形成のために重要であることが分かったが、シナプスが、どうして軸索と樹状突起の間だけで特異的に形成されるのか、その機構が未解明であった。研究を進めた結果、軸索と樹状突起のそれぞれに分布するネクチン1とネクチン3(免疫グロブリンスーパーファミリー分子)の間のヘテロフリックな結合がまず認識分子として働き、これにカドヘリンが相互作用してシナプス結合を強化する過程が明らかになった。
カドヘリンがシナプス形成のために重要であることが分かったが、シナプスが、どうして軸索と樹状突起の間だけで特異的に形成されるのか、その機構が未解明であった。研究を進めた結果、軸索と樹状突起のそれぞれに分布するネクチン1とネクチン3(免疫グロブリンスーパーファミリー分子)の間のヘテロフリックな結合がまず認識分子として働き、これにカドヘリンが相互作用してシナプス結合を強化する過程が明らかになった。 シナプス形成に関与するカドヘリン-カテニン複合体の新しい機能を明らかにすることができた。NMDA受容体を刺激すると、カルパインが活性化することが知られるが、このタンパク分解酵素がβ-カテニンのN末端を切断し安定化すること、そして安定化β-カテニンは核に入り転写制御に関わることを示した。これは、生理的刺激がβ-カテニン依存の転写制御系を活性化することを明らかにした初めての発見である。
シナプス形成に関与するカドヘリン-カテニン複合体の新しい機能を明らかにすることができた。NMDA受容体を刺激すると、カルパインが活性化することが知られるが、このタンパク分解酵素がβ-カテニンのN末端を切断し安定化すること、そして安定化β-カテニンは核に入り転写制御に関わることを示した。これは、生理的刺激がβ-カテニン依存の転写制御系を活性化することを明らかにした初めての発見である。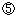 上記以外にも、カドヘリン系が網膜の水平細胞のシナプス形成のために必須であることなどについて明らかにすることができた。
上記以外にも、カドヘリン系が網膜の水平細胞のシナプス形成のために必須であることなどについて明らかにすることができた。
2)カドヘリンスーパーファミリー分子Flamingoの細胞極性形成における役割
特別推進研究の初期に、新しいカドヘリンスーパーファミリーFlamingoをショウジョウバエを用いて同定し、平面内細胞極性形成の制御因子であることを明らかにした。その後、この分子の脊椎動物ホモログについて研究を進め、たとえば小脳プルキンエ細胞樹状突起のパターンニングに重要な役割を果たすことを明らかにし、動物界を通して細胞の形態制御に関わることを示すことができた。
3)カドヘリン接着装置機構に関する細胞生物学的研究の成果
特別推進研究の期間中に、カドヘリン結合因子の一つp120カテニンの研究を開始した。その後、この分子の研究は2つの方向に発展した。一つは、この分子が微少管に結合することの発見。これにより、カドヘリン接着装置と微少関係が相互作用するという、従来あまり重要視されてこなかった問題が浮かびあがり、これに関する研究は現在大きく発展しており、その機構、生物学的意義の解明につながっている(論文投稿中)。もう一つについては、p120カテニンに結合するタンパクのスクリーニングをした結果、Cdc42に特異的なGEF、Tubaが見つかり、その機能の研究に発展した。Tubaは、細胞結合領域のアクチン重合を制御して、細胞境界の張力を制御することが明らかになった。さらに、これらの細胞生物学的研究の延長として、予期せぬ現象“cadherin flow”を発見することができた。
4)網膜幹細胞維持におけるWntシグナル分子の役割
特別推進研究期間の後半に、解離した網膜細胞が再集合するとき、他の細胞のコンディションメディウムを加えておくと、再集合体の中での組織形成が著しく促進されることが分かった。有効因子を同定することは、神経組織形成機構を理解する上で重要と考え、研究を進めた結果、Wntが有効であること、さらに、網膜の幹細胞が分布する辺縁領域にWnt2bが発現していることが分かった。これらの現象を詳細に調べた結果、Wnt2bが網膜幹細胞の維持のために必要であること、この作用が解離細胞からの網膜組織形成のために働くことなどを明らかにした。研究の動機とは異なる方向に発展したが、幹細胞の維持機構の解明という重要な貢献をすることができた。
2.特別推進研究の研究成果が他の研究者による活用された状況はどうか
(1)学界への貢献の状況
1)シナプス形成におけるカドヘリン接着装置の役割
シナプス結合の形成にカドヘリンが関与することを実験的に証明したのは我々が始めてである。その後、この問題の重要性に対する認識は、専門家の間に次第に広まり、今や、ほぼ教科書的スタンダードになりつつある。
2)カドヘリンスーパーファミリー分子Flamingoの細胞極性形成における役割
平面内極性形成機構の研究は、近年の形態形成学における主要テーマの一つである。Frizzled,Disheveledなどの関与が知られていたが、Flamingoが重要であるとの発見は、平面内極性形成機構の解明のために新たな展開をもたらした。とりわけ、これらの成分が、細胞極性に対応して局所的分布をするという発見は大きい。以降の研究では、本発見に影響され、Frizzled, Disheveled等も同様な局在することが気づかれ、今や本分野の研究は、これらの分子間の相互作用の結果生じるシグナル機構の解明に注がれている。
3)カドヘリン接着装置機構に関する細胞生物学的研究の成果
カドヘリンによる細胞間接着装置とはどのようなもので、また、他の細胞内システムとどのように関係しながら働いているかについて、いくつかの重要な知見を蓄積できた。とりわけcadherin flowの発見は、細胞間接着が“静的な”分子間相互作用によっておきるという先入観を覆し、カドヘリン分子が極性をもって流動する状態でおきている場合があることを明らかにしたもので、この意外性は本研究分野に大きなインパクトを与えた。また、最近、カドヘリンはカテニンを介してアクチン繊維と結合しているとする従来のモデルは正しくないという見解が発表された。しかし私達は、カテニンとアクチンは、EPLINによって繋ぎ止まれているということを発見し、この問題に対して正しい解を提示することができた。
4)網膜幹細胞維持におけるWntシグナル分子の役割
幹細胞の生成・維持機構に対する関心は、再生医療の実現に向けて、近年、著しく高まってきている。この分野がまだ未成熟な中、いち早くWntシグナル系の関与を示すことができたことは大きい。この成果は、網膜視細胞変性症の再生医療学的治療を目指す理研・網膜再生医療研究チームによって直ちに応用され、網膜に人為的に傷害を与えたマウスモデル実験系においてWntの投与が視細胞の再生を促進することが明らかにされた(J. Neurosci. 27:4210-4219, 2007)。
(2)論文の引用状況
調査日 2007年11月15日
研究期間中に発表された論文
- Yagi, T. and Takeichi, M. (2000) Cadherin superfamily genes: functions, genomic organization, and neurologic diversity. Genes & Dev. 14, 1169-1180. 「カドヘリンスーパーファミリーの機能、および、神経疾患との関係についての総説」 268件
- Niwa, R., Nagata-Ohashi, K., Takeichi, M., Mizuno, K., and Uemura, T. (2002) Control of actin reorganization by Slingshot, a family of phosphatases that dephosphorylate ADF/cofilin. Cell 108, 233-246. 「Slingshotと命名した脱リン酸化酵素がコフィリンの活性制御を介してアクチンの再編成のために働くことを明らかにした」 159件
- Usui, T., Shima, Y., Shimada, Y., Hirano, S., Burgess, R.W., Schwarz, T.L., Takeichi, M., and Uemura, T. (1999) Flamingo, a seven-pass transmembrane cadherin, regulates planar cell polarity under the control of Frizzled. Cell 98, 585-595. 「カドヘリンスーパーファミリーに属する“Flamingo”の一次構造、および、この分子が平面内細胞極性に必要であることを示した論文」 231件
- Togashi, H., Abe, K., Mizoguchi, A., Chisaka, O., and Takeichi, M. (2002) Cadherin regulates dendritic spine morphogenesis. Neuron 35, 77-89. 「カドヘリンが興奮性神経細胞樹状突起上のスパインの形態形成に重要であることを初めて明らかにした論文」 157件
- Aono, S., Nakagawa, S., Reynolds, A.B., and Takeichi, M. (1999) p120ctn acts as an inhibitory regulator of cadherin function in colon carcinoma cells. J. Cell Biol. 145, 551-562. 「カドヘリン結合分子p120カテニンが、カドヘリン活性の抑制因子として働くことを見出した論文」 150件
- Nakagawa, S., and Takeichi, M. (1998) Neural crest emigration from the neural tube depends on regulated cadherin expression. Development 125, 2963-2971. 「神経冠細胞の神経管からの遊走が、カドヘリンの発現制御に依存していることを示した論文」 133件
- Shibamoto. S., Higano, K., Takada, R., Ito, F., Takeichi, M., and Takada, S. (1998) Cytoskeletal reorganization by soluble Wnt-3a protein signalling. Genes to Cells, 3:659-70. 「Wnt-3aが細胞骨格の再構成に作用することを示した論文。この研究によって、Wnt-3aの分泌細胞系が確立され、以降、本系または同様な実験系がWnt研究のために世界中で使用されるようになった」 127件
- Watabe-Uchida, M., Uchida, N., Imamura, Y., Nagafuchi, A., Fujimoto, K., Uemura, T., Vermeulen, S., van Roy, F., Adamson, E.D., and Takeichi, M. (1998) α-Catenin-vinculin interaction functions to organize the apical junctional complex in epithelial cells. J. Cell Biol. 142, 847-857. 「カドヘリン結合因子の1つα-cateninの活性として、ヴィンキュリンとの結合が見出され、これが上皮細胞における頂端部の接着構造形成のために働くことを明らかにした論文」 118件
- Shimada, Y., Usui, T., Yanagawa, S., Takeichi, M., and Uemura, T. (2001) Asymmetric colocalization of Flamingo, a seven-pass transmembrane cadherin, and Dishevelled in planar cell polarization. Current Biology 11, 859-863. 「平面内極性を制御することが知られる3つのタンパク質、Flamingo, Dishevelled, Frizzledがシグナル複合体を形成しており、その細胞内局在が細胞骨格系の再編成を統括することを示唆した論文」 69件
- Inoue, T., Tanaka, T., Takeichi, M., Chisaka, O., Nakamura, S., and Osumi, N. (2001) Role of cadherins in maintaining the compartment boundary between the cortex and striatum during development. Development 128, 561-569. 「大脳皮質と線条体の境界が、複数のカドヘリンの発現パターンによって決められることを示唆した論文」 69件
研究期間終了後に発表された論文
- Kubo, F., Takeichi, M., and Nakagawa, S. (2003) Wnt2b controls retinal cell differentiation at the ciliary marginal zone. Development 130,587-598. 「Wnt2bが網膜細胞の未分化状態を維持することに働いていることを示す論文」 76件
- Abe, K., Chisaka, O., van Roy, F., and Takeichi, M. (2004) Stability of dendritic spines and synaptic contacts is controlled by αN-catenin. Nature Neuroscience 7, 357-363. 「αN-cateninが、シナプス構造の安定性を維持するために必要であることを明らかにした論文」 55件
- Takeichi, M., and Abe, K. (2005) Synaptic contact dynamics controlled by cadherin and catenins. Trends in Cell Biol., 15, 216-221. 「特別推進研究を通じて明らかになった、カドヘリンとカテニンによるシナプス制御機構をまとめた総説」 34件
- Shima, Y., Kengaku, M., Hirano, T., Takeichi, M., and Uemura T. (2004) Regulation of dendritic maintenance and growth by a mammalian 7-pass transmembrane cadherin. Develop. Cell 7, 205-216. 「脊椎動物のFlamingoホモログがプルキンエ細胞樹状突起のパターン形成に関与することを初めて示した論文」 27件
- Tanoue, T., and Takeichi, M. (2004) Mammalian Fat1 cadherin regulates actin dynamics and cell-cell contact. J. Cell Biol. 165, 517-528. 「脊椎動物Fatカドヘリンホモログがアクチン重合と細胞間接着に関与することを初めて明らかにした論文」 27件
- Nakagawa, S., Takada, S., Takada, R., and Takeichi, M. (2003) Identification of the laminar-inducing factor: Wnt-signal from the anterior rim induces correct laminar formation of the neural retina in vitro. Develop. Biol., 260, 414-425. 「Wnt2bが、解離した網膜細胞の再構成に重要な役割を果たすことを明らかにした論文」 24件
- Kubo, F., Takeichi, M., and Nakagawa, S. (2005) Wnt2b inhibits differentiation of retinal progenitor cells in the absence of Notch activity by downregulating the expression of proneural genes. Development 132, 2759-2770. 「Wnt2bとNotchは別の経路で網膜細胞分化を抑制することを見出した論文」19件
- Tanoue, T., and Takeichi, M. (2005) New insights into Fat cadherins. J. Cell Sci., 118, 2347-2353. 「我々が明らかにした脊椎動物Fat カドヘリンの機能を中心にこの分子に関する最近の知見をまとめた総説」 16件
- Abe, K., and Takeichi, M. (2007) NMDA Receptor activation induces calpain-mediated β-catenin cleavages for triggering gene expression. Neuron 53, 387-397. 「グルタミン酸によるNMDA Receptorの活性化が、カルパインの活性化を介してβ-cateninのN末端部を切断・安定化し、その転写制御活性を誘発することを明らかにした画期的論文」6件
- Togashi, H., Miyoshi, J., Honda, T., Sakisaka, T., Takai, T., and Takeichi, M. (2006) Interneurite affinity is regulated by heterophilic nectin interactions in concert with the cadherin machinery. J. Cell Biol., 174, 141-151. 「シナプス形成時における軸索と樹状突起間の認識は、カドヘリンとネクチン分子群の協調作業によって起きることを示した論文」6件
3.その他、効果・効用等の評価に関する情報
(1)研究成果の社会への還元の状況
本研究は純粋な基礎研究であるため、特許取得の申請等はしていない。得られた成果の多くは、当面は、生命現象のしくみのより深い理解という形で社会に還元する。その方法については、理化学研究所及び当研究所発生・再生科学総合研究センターから出版される種々の広報誌、さらには、一般市民向けの講演会を通じて行っている。Wnt2bが網膜幹細胞の維持に関与することの発見は、発生・再生科学総合研究センター網膜再生医療研究グループ(高橋政代チームリーダー)により発展させられ、網膜視細胞変性症の治療に貢献できる可能性が示唆された。
(2)研究計画に関与した若手研究者の成長の状況
本研究に参画した若手研究者は、助手2名(研究開始の段階で分担研究者として1名、途中から1名)、及び、大学院生(15名)である。現在のポジションについて列記する(括弧内は特別推進研究期間中のポジション)。
- 上村匡
 助手)
助手) - 京都大学大学院生命科学研究科 教授
- 中川真一助手)
- 理化学研究所独立主幹研究員
- 青野真也大学院生)
- 京都大学再生医科学研究所 助教
- 本庶仁子大学院生)
- オレゴン大学神経科学研究所 博士研究員
- 碓井理夫大学院生)
- 京都大学大学院生命科学研究科 助教
- 鈴木祥宏大学院生)
- 理化学研究所 研究員
- 堀川一樹大学院生)
- 北海道大学電子科学研究所 特任準教授
- 島康彦大学院生)
- ブランダイス大学 博士研究員
- 島田裕子大学院生)
- エール大学医学部遺伝学教室 博士研究員
- 一居哲夫大学院生)
- 大阪大学JST-ERATO(エラート) 研究員
- 大谷哲久大学院生)
- 理化学研究所 研究員
- 富樫英大学院生)
- 神戸大学大学院医学研究科 助教
- 田辺光志大学院生)
- 理化学研究所 研究員
- 亀谷祥子大学院生)
- カリフォルニア大学サンフランシスコ校 博士研究員
- 門脇正和大学院生)
- シスメックス株式会社 社員
- 安部健太郎大学院生)
- 京都大学大学院生命科学研究科 助教
- 久保郁大学院生)
- 理化学研究所基礎科学特別研究員
-- 登録:平成21年以前 --